Функции ретикулярной формации ствола мозга. Функции ретикулярной формации. Особенности структурно-функциональной организации. Функции нисходящей ретикулярной системы
Сложное строение человеческого мозга открывает тайны нашего поведения, объясняет законы мыслительной деятельности, протекание эмоций и чувств. Каждое полушарие головного мозга отвечает за свои специфические функции и задачи (например, известно, что правое отвечает за логику, а левое - за воображение и фантазию), но имеются также структуры, которые обеспечивают единую и согласованную работу всей центральной нервной системы. Одной из таких структур является ретикулярная формация.
Общие сведения
Ретикулярная формация - это участок стволового отдела мозга, представленный разветвленной сетью нервных клеток и ядер, соединяющих между собой различные отделы головного мозга. В отличие от других структур, - например, таламуса, гипоталамуса, мозжечка, - которые имеют некую цельную форму (ядра, железы), ретикулярная формация не представлена единым морфологическим образованием, а является «сетью» (от латинского reticulum — сеть) дендритов и аксонов, которые с разной степенью плотности проникают между отделами и структурами головного мозга, объединяя их между собой и обеспечивая их совместную деятельность.
Метафорически выражаясь: если наш мозг представить в виде некоего изделия, скажем, рубашки, то ретикулярная формация - это нити, которыми рубашка сшита. Ретикулярная формация пронизывает структуры продолговатого, среднего мозга и моста, имеет непосредственные связи с мозжечком, спинным мозгом, таламусом и опосредованные - с вышележащими отделами: гипоталамусом, зрительными ядрами и корой.
Как устроена
В состав ретикулярной формации входит огромное количество нейронов с разветвленными дендритами и длинными аксонами, за счет чего становится возможным передавать нервные импульсы в различные отделы головного и спинного мозга. При этом можно выделить две наиболее крупные группы нейронных скоплений:
- Ретикулотегментальное ядро, нейроны которого получают сигналы от вышележащих отделов ГМ (четверохолмие, таламус) и передают их далее в структуры мозжечка, регулируя тем самым некоторые жизненно важные двигательные функции: координация взгляда, движения глаз.
- Латеральное ядро, нейроны которого восходят от структур спинного мозга и вестибулярных ядер и обеспечивают информирование коры ГМ о положении тела в пространстве, участвуют в регуляции дыхания и сосудистой иннервации.
- Кроме того, в состав ретикулярной формации входят нейроны, которые принимают важное участие в работе центров терморегуляции, насыщения и голода.

Основные функции
Основное предназначение ретикулярной формации состоит в сенсорном анализе многочисленных сигналов, поступающих от различных отделов ГМ.
За счет тесных связей со спинным мозгом она также принимает самое активное участие в моторной регуляции, начиная от глотательного рефлекса и заканчивая сложными двигательными операциями. Кроме того, ретикулярная формация осуществляет активирующее воздействие на весь ГМ в целом, участвуя в регуляции циклов сна и бодрствования.
В общем виде функции ретикулярной формации сводятся к следующим:
- Регуляция скелетной мускулатуры (участвует в управлении движениями тела) и вегетативных функций (дыхание, чихание, кровообращение и т.п.).
- Контроль процессов сна и бодрствования (за счет оказания активирующего и тормозящего воздействия на кору больших полушарий).
- Активизирующая функция (проявляется в том, что ретикулярная формация обеспечивает постоянное тоническое возбуждение коры ГМ, за счет чего становится возможным поддержание внимания, сознания и протекание мыслительных процессов.)
- Обработка сигналов, поступающих из внешней и внутренней среды.
Отличительные особенности работы ретикулярной формации связаны, прежде всего, с определенными свойствами ее :
Нервные клетки, входящие в состав ретикулярной формации, обладают повышенной способностью к тоническому возбуждению. Это означает, что бо́льшая часть нейронов находятся в постоянном возбуждении и генерирует нервные импульсы, которые передаются в вышележащие отделы ГМ. Такая тоническая активность обусловлена рядом факторов:
- Проходимость большого количества сигналов через структуры ретикулярной формации. Приведем простую аналогию: представьте себе рояль или какой-либо другой струнный инструмент. Понятно, что когда мы напрямую касаемся струн, они начинают колебаться и издавать звук. То же самое происходит и с нервными клетками, когда к ним приходят сигналы от других нейронов.Однако представим далее, что мы не касаемся непосредственно струн инструмента, но, скажем, скачем рядом с ним, сильно стуча ногами о пол. Звука инструмента мы, может быть, и не услышим, но едва заметное колебание струн все же будет иметь место. То же самое происходит и с нейронами ретикулярной формации. Поскольку через нее постоянно проходят какие-то сигналы (как афферентные, так и эфферентные) от различных структур ЦНС, то это и создает постоянное тоническое возбуждение нейронов ретикулярной формации, за счет того, что она находится в эпицентре постоянного нервно-импульсного обмена.
- Повышенная чувствительность нейронов к химическим веществам (гормоны, медикаменты, психотропные вещества). Чашка кофе, выпитая утром, «включает» именно структуры ретикулярной формации и за счет длительного сохранения возбуждения в ее нейронах поддерживает нас в активном состоянии.
Нисходящее и восходящее влияние РФ
Как уже отмечалось, ретикулярная формация оказывает возбуждающее и тормозящее действие на различные отделы ГМ. При этом можно выделить два отдела, которые специализируются на передаче возбуждения в те или иные структуры мозга.
Нисходящий отдел: представлен вегетативными и двигательными центрами и оказывает нисходящее влияние на отделы спинного мозга. Соответствующие нейронные скопления регулируют деятельность дыхательного, сосудодвигательного, слюноотделительного центров, а также центров, отвечающих за построение простых и сложных двигательных реакций. Это указывает на решающую роль именно центральной нервной системы в регуляции даже элементарных безусловных рефлексов. Стимуляция нисходящего отдела приводит к затормаживанию спинальных центров и вызывает в естественной среде состояние глубоко сна (спать «без задних ног»). Тот же эффект можно вызвать и искусственным путем, например, вводя человека в состояние транса или наркоза.
Восходящий отдел: представлен нервными волокнами, соединяющими структуры ретикулярной формации с вышележащими отделами: таламусом, гипоталамусом, мозжечком и корой. Восходящее влияние оказывает стимулирующее действие на кортикальные структуры и обеспечивает активное состояние сознания. Восходящее влияние не прекращается, даже когда мы спим. Если бы наш мозг мог полностью «отключаться», то каждое пробуждение было бы сродни рождению: кто я? Где я? Как я сюда попал? Однако за счет работы ретикулярных структур у нас сохраняется возможность всегда возвращаться в то исходное состояние сознания, в котором мы находились до момента сна. Кроме того, и во время ночного отдыха у нас остается способность реагировать на некоторые жизненно важные раздражители, т.е. мы, как правило, не спим «мертвым сном» и можем проснуться, если рядом зашевелился и заплакал ребенок, что-то громко упало и т.п.
Проявление повреждения структур
Ретикулярная формация играет значимую роль в интегративной деятельности всего головного мозга. За счет выполнения функции ведущего проводника всех видов нервных импульсов во все отделы ЦНС, ретикулярная формация находится в постоянной работе. Чрезмерные умственные и эмоциональные перегрузки вредны для мозга в целом и для ретикулярной формации, в частности. К счастью, своевременный прием седативных препаратов может (за счет повышенной восприимчивости нейронов к химическому воздействию) быстро исправить ситуацию и нормализовать состояние.
Однако возможны и менее благоприятные исходы. Повреждение возможно в результате черепно-мозговых травм, онкологических заболеваний головного мозга, инфекционных поражений.
Основным проявлением неблагополучия является потеря сознания.
Нарушение восходящих связей обнаруживает себя в состоянии апатии, слабости, повышенной сонливости, двигательной растороможенности, нарушениях ночного сна. Нередки сопутствующие вегетативные расстройства.
Ретикулярная формация - это комплекс нейронов ствола головного мозга и частично спинного мозга, который имеет обширные связи с различными нервными центрами, корой головного мозга и между собой.
Ретикулярная формация - это формация, идущая от спинного мозга к таламусу в ростральном (к коре) направлении. Кроме участия в обработке сенсорной информации, ретикулярная формация оказывает активизирующее воздействие на кору головного мозга, контролируя таким образом деятельность спинного мозга. С помощью данного механизма осуществляется контроль тонуса скелетной мускулатуры, половой и вегетативных функций человека. Впервые механизм воздействия ретикулярной формации на мышечный тонус был установлен Р.Гранитом (R.Granit): он показал, что ретикулярная формация способна изменять активность γ-мотонейронов, в результате чего их аксоны (γ-эфференты) вызывают сокращение мышечных веретён, и, как следствие, усиление афферентной импульсции от мышечных рецепторов. Эти импульсы, поступая в спинной мозг, вызывают возбуждение α-мотонейронов, что и является причиной тонуса мышц.
Установлено, что участие в выполнении данной функции ретикулярной формации принимают два скопления нейронов: нейроны ретикулярной формации моста и нейроны ретикулярной формации продолговатого мозга. Поведение нейронов ретикулярной формации продолговатого мозга схоже с поведением нейронов ретикулярной формации моста: они вызввают активацию α-мотонейронов мышц-сгибателей и, следовательно, тормозят активность α-мотонейронов мышц-разгибателей. Нейроны ретикулярной формации моста действуют ровно наоборот, возбуждают α-мотонейроны мышц-разгибателей и тормозят активность α-мотонейронов мышц-сгибателей. Реткулярная формация имеет связь с мозжечком (часть информации от него идёт к нейронам продолговатого мозга (от ядер пробковидного и шаровидного мозжечка), а от шатра - к нейронам моста) и с корой головного мозга, от которой получает информацию. Это позволяет утверждать, что ретикулярная формация является коллектором неспецеффического сенсорного потока, возможно участвующим в регуляции мышечной активности. Хотя пока необходимость в ретикулярной формации, дублирующей функции нейронов вестибулярных ядер и красного ядра, остаётся непонятной.
Ретикулярная формация представлена рассеянными клетками в покрышке ствола мозга и в спинном мозге. Ряд клеток ретикулярной формации в стволе мозга являются жизненно важными центрами:
1. дыхательным (центр вдоха и выдоха) - в продолговатом мозге;
2. сосудодвигательным - в продолговатом мозге;
3. центром координации взора (ядра Кахаля и Даркшевича) - в среднем мозге;
4. центром терморегуляций - в промежуточном мозге;
5.
центром
голода и насыщения - в промежуточном мозге.
6. Ретикулярная формация выполняет следующие функции:
Обеспечение сегментарных рефлексов: рассеянные клетки являются вставочными нейронами спинного мозга и ствола головного мозга (рефлекс глотания);
Поддержание тонуса скелетной мускулатуры: клетки ядер ретикулярной формации посылают тонические импульсы на двигательные ядра черепных нервов и двигательные ядра передних рогов спинного мозга;
Обеспечение тонической активности ядер ствола головного мозга и коры полушарий, что необходимо для дальнейшего проведения и анализа нервных импульсов;
Коррекция при проведении нервных импульсов: благодаря ретикулярной формации импульсы могут либо существенно усиливаться, либо существенно ослабляться в зависимости от состояния нервной системы;
Активное влияние на высшие центры коры больших полушарий, что приводит к либо снижению тонуса коры, апатии и наступлению сна, либо к повышению работоспособности, эйфории;
Участие в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и других вегетативных функций (центры ствола мозга);
Участие в регуляции сна и бодрствования: голубое пятно, ядра шва - проецируются на ромбовидную ямку;
Обеспечение
сочетанного поворота головы и глаз: ядра Кахаля и
Даркшевича.
Основным нисходящим трактом ретикулярной формации является ретикулоспинальный, который проходит по стволу к нейронам двигательных ядер передних рогов спинного мозга и двигательных ядер черепных нервов, а также к вставочным нейронам вегетативной нервной системы.
От ретикулярных ядер зрительного бугра к различным областям коры больших полушарий идут таламо-корковые волокна: они заканчиваются во всех слоях коры большого мозга, осуществляя активацию коры, необходимую для восприятия специфических раздражений.
Исследования влияния ретикулярной формации на поведениеМикроскопические электроды вживляли животному в клетки ретикулярной формации. Когда оно засыпало, эти области центральной нервной системы раздражали слабым током и одновременно с помощью электроэнцефалографа записывали электрическую активность мозга. Животное пробуждалось немедленно, а на электроэнцефалограмме появлялись быстрые и частые колебания, характерные для бодрствующего мозга. Причем эти изменения наблюдались во всех областях коры больших полушарий.
В другом эксперименте разрушали определенные участки ретикулярной формации. В результате поведение животного резко менялось. Оно погружалось в спячку, а электроэнцефалограмма регистрировала медленные, «дремотные» электрические волны. Вывести животное из состояния сна, применяя даже очень сильные внешние раздражители, как правило, не удавалось.
Важный вывод сделали ученые: ретикулярная формация оказывает активизирующее влияние на кору больших полушарий головного мозга. Она является своеобразным «энергетическим центром» мозга, без которого нервные клетки коры, различные ее отделы, весь мозг в целом не могут выполнять свои сложные многообразные функции. Она принимает непосредственное участие в процессах регулирования не только сна, но и бодрствования.
Экспериментальные работы физиологов позволили дать объяснения наблюдениям хирургов. Во время операций на мозге можно делать разрезы коры больших полушарий, удалять часть мозговой ткани, и человек не потеряет сознания. Но стоит скальпелем задеть ретикулярную формацию, как больной впадает в глубокий сон.
Каким же образом осуществляется активизирующее влияние ретикулярной формации на мозг? Откуда она черпает энергию для поддержания рабочего состояния коры больших полушарий, определяя тем самым бодрствование организма?
В настоящее время хорошо изучены так называемые специфические нервные пути, по которым в мозг поступает информация от органов чувств. Именно таким образом кора мозга «узнает» о характере действующего на организм раздражителя и в соответствии с этим посылает сигналы к различным органам и системам.
Исследования ретикулярной формации показали, что от всех без исключения чувствительных неявных волокон, направляющихся с периферии в кору больших полушарий, отходят ответвления, заканчивающиеся на поверхности клеток ретикулярной формации. Любое внешнее раздражение - световое, звуковое, болевое, тактильное (осязательное) - возбуждает ретикулярную формацию. Она в этот момент как бы «заряжается» энергией. И, в свою очередь, как «энергетический центр» мозга, определяет уровень работоспособности коры больших полушарий.
Активизируя все отделы мозга, ретикулярная формация обеспечивает точный анализ и синтез многообразной информации, поступающей из внешнего мира в кору больших полушарий по специфическим нервным путям. В этом отношении очень показателен такой эксперимент. Обезьяны, которых обучили выбирать один из двух быстросменяющихся предметов, делали это значительно скорее и точнее, если одновременно с помощью вживленных электродов раздражалась ретикулярная формация.
И еще одно важное наблюдение сделали физиологи. Оказалось, что ретикулярная формация очень тонко реагирует не только на нервные сигналы, но и на растворенные в крови физиологически активные вещества: сахар, кислород, углекислоту, гормоны. Среди них наибольшее значение в поддержании активности ретикулярной формации принадлежит адреналину - гормону надпочечников.
При эмоциональном перенапряжении, состояниях аффекта - гневе, ярости, страхе - происходит длительное возбуждение ретикулярной формации. Это возбуждение поддерживает адреналин, который усиленно выделяется в кровь.
Активность ретикулярной системы во многом определяется и другими химическими веществами, содержание которых в крови выше или ниже определенного критического уровня может стать роковым для организма. Это прежде всего насыщенность крови кислородом и углекислым газом. Например, если у спящего человека затруднено дыхание, то в крови начинает накапливаться углекислота. Она возбуждает ретикулярную формацию, и человек просыпается.
Дальнейшее изучение деятельности ретикулярной формации показало, что она не автономна, не независима, а находится под постоянным контролем коры больших полушарий. При этом уровень функциональной активности ретикулярной формации тем выше, чем ниже возбуждение коры мозга. Так, например, снижение функциональной активности коры больших полушарий или их удаление в опытах на животных приводит к значительному возбуждению ретикулярной формации. Поведение животных резко меняется, они становятся агрессивными.
Клинические наблюдения и полученные в физиологических лабораториях экспериментальные данные показали также, что ретикулярная формация имеет прямое отношение к формированию эмоций.
Исследования строения и функций ретикулярной формации нашли широкое применение в клинической практике, в нейро- и психофармакологии. Оказалось, что апатия, вялость, сонливость и, напротив, бессонница, раздражительность могут возникать в связи с расстройством деятельности ретикулярной формации. Определенную роль она играет в возникновении многих заболеваний центральной нервной системы.
Поскольку клетки ретикулярной формации необычайно чувствительны к растворенным в крови химическим веществам, то значит, что с помощью лекарственных средств можно регулировать деятельность клеток - повышать или, наоборот, подавлять их возбудимость.
Ретикулярная формация ствола мозга представляет собой комплекс нейронов, имеющих обширные связи с разными нервными центрами, друг с другом и корой полушарий. Она пролегает в ростральном направлении к таламусу. Рассмотрим далее ее особенности.
Функции ретикулярной формации
В задачи комплекса входит обработка сенсорной информации. Кроме этого, ретикулярная формация обеспечивает активизирующее воздействие на кору, осуществляя контроль деятельности спинного мозга. За счет этого регулируются тонус скелетных мышц, работа вегетативной и половой систем человека.
Механизм действия
Впервые он был выявлен Р. Гранитом. Ученый установил, что может влиять на активность γ-мотонейронов. Вследствие этого γ-эфференты (их аксоны) провоцируют сокращение веретен мускулатуры и, соответственно, повышение афферентной импульсации мышечных рецепторов. Поступающие в спинной мозг сигналы провоцируют возбуждение α-мотонейронов. Это и обуславливает тонус мускулатуры. Было установлено, что в реализации этой функции участвуют нейроны формации моста и продолговатого мозга. Их поведение диаметрально противоположно. Последние провоцируют активацию α-мотонейронов в мышцах-сгибателях и, соответственно, тормозят их в разгибателях. Нейроны моста действуют наоборот. Ретикулярная формация связана с мозжечком и корой, от которой поступает информация. Это позволяет сделать вывод, что она выступает в качестве коллектора неспецифического сенсорного потока, который, возможно, участвует в регуляции активности мускулатуры. Однако в настоящее время еще не выяснена необходимость формации, дублирующей задачи нейронов в красном и вестибулярных ядрах.

Структура
Ретикулярная формация образуется рассеянными клетками. Некоторые из них считаются жизненно важными образованиями. В частности, можно выделить центры:
- Дыхательный и сосудодвигательный. Они располагаются в продолговатом мозге.
- Координации взора. Он находится в среднем мозге.
- Голода, насыщения и терморегуляции. Они располагаются в промежуточном мозге.
В качестве ключевого тракта выступает ретикулоспинальный. Он проходит к нейронам в двигательных ядрах передних спинномозговых рогов и черепных нервов по стволу и к вставочным элементам нервной вегетативной системы. От них пролегают таламо-корковые волокна. Они обеспечивают активацию коры, которая необходима для восприятия специфических раздражителей. Эти таламо-корковые волокна заканчиваются во всех корковых слоях.

Научные наблюдения
В ходе исследований было выявлено, что ретикулярная формация обладает активизирующим воздействием на кору. Этот нейронный комплекс выступает в качестве своеобразного "энергетического центра". Без него нервные клетки коры, разные ее отделы, а также весь мозг в целом не смогут выполнять все свои многообразные сложные задачи. Комплекс нейронов непосредственно участвует в процессе регулирования сна и бодрствования. Результаты экспериментов позволили объяснить некоторые наблюдения хирургов. Так, в процессе операций на мозге могут быть сделаны разрезы в коре полушарий, удалена часть ткани. При этом пациент сознания не потеряет. Однако, если скальпелем будет задета , человек впадет в глубокий сон.
Специфика работы
Сегодня достаточно хорошо изучены специфические нервные каналы, по которым от органов чувств передается информация в мозг. Именно так кора узнает о характере раздражителя, действующего на организм. В соответствии с этим она посылает разные импульсы к системам и органам. Исследования показали, что от всех волокон, направленных от периферии к коре, отходят ответвления. Они заканчиваются на поверхности клеток формации. Внешнее раздражение любого характера оказывает на нее возбуждающее действие. В этот момент происходит своего рода "зарядка энергией". Выступая как мозговой центр, формация определяет степень работоспособности коры. Активизируя все отделы, она обеспечивает точный синтез и анализ многообразия информации, которая поступает в кору из внешнего мира.

Реакция на вещества организма
Ретикулярная формация чувствительна не только к нервным сигналам, но и к растворенным в крови соединениям. В частности, речь о сахаре, гормонах, углекислоте, кислороде. Особое значение среди этих веществ имеет адреналин. При эмоциональном перенапряжении - при гневе, страхе, состоянии аффекта, ярости - отмечается продолжительное возбуждение формации. Его поддерживает адреналин, усиленно выделяющийся в кровь. Активность комплекса во многом определяют и другие химические соединения. В первую очередь это углекислый газ и кислород. К примеру, если у человека во сне затруднено дыхание, то СО 2 начинает накапливаться в крови. Углекислота активизирует ретикулярную формацию, вследствие чего человек просыпается.

Заключение
Клинические исследования и экспериментальные данные, полученные в физиологических лабораториях, показали, что ретикулярная формация прямо связана с возникновением эмоций. Итоги изучения ее строения и задач, которые она реализует, широко применяются в психо- и нейрофармакологии. Было установлено, что вялость, апатия, сонливость или раздражительность, бессонница могут обуславливаться расстройством в работе ретикулярной формации. Этот нейронный комплекс также выполняет определенную роль в процессе возникновения многих патологий ЦНС.
1. Анатомическое строение и волоконный состав…………………………3
2. Неспецифические нисходящие влияния………………………………….3
3. Восходящие влияния………………………………………………………5
4. Свойства ретикулярных нейронов………………………………………..8
5. Выводы……………………………………………………………………..10
Список литературы…………………………………………………………………...12
1. Анатомическое строение и волоконный состав
Ретикулярная формация - совокупность нейронов и соединяющих их нервных волокон, расположенных в стволе мозга и образующих сеть.
Ретикулярная формация простирается через весь ствол головного мозга от верхних шейных спинальных сегментов до промежуточного мозга. Анатомически она может быть разделена на ретикулярную формацию продолговатого мозга, варолиевого моста и среднего мозга. Вместе с тем, в функциональном отношении в ретикулярной формации разных отделов мозгового ствола есть много общего. Поэтому целесообразно рассматривать ее как единую структуру.
Ретикулярная формация представляет собой сложное скопление нервных клеток, характеризующихся обширно разветвленным дендритным деревом и длинными аксонами, часть из которых имеет нисходящее направление и образует ретикулоспинальные пути, а часть восходящее. В ретикулярную формацию поступает большое количество путей из других мозговых структур. С одной стороны, это коллатерали волокон, проходящих через ствол мозга сенсорных восходящих систем, эти коллатерали заканчиваются синапсами на дендритах и соме нейронов ретикулярной формации. С другой стороны, нисходящие пути, идущие из передних отделов мозга (в том числе, пирамидный путь), тоже дают большое количество коллатералей, которые входят в ретикулярную формацию и вступают в синаптические соединения с ее нейронами. Обилие волокон поступает к нейронам ретикулярной формации из мозжечка. Таким образом, по организации своих афферентных связей эта система приспособлена к объединению влияний из различных мозговых структур. Выходящие из нее пути могут оказывать в свою очередь влияния как на вышележащие, так и на нижележащие мозговые центры.
Нейронная организация ретикулярной формации до сих пор остается недостаточно изученной. В связи с чрезвычайно сложным переплетением в ней отростков различных клеток разобраться в характере межнейронных связей этой области очень трудно. Вначале широко распространенным было представление, что отдельные нейроны ретикулярной формации тесно связаны друг с другом и образуют нечто подобное нейропилю, в котором возбуждение распространяется диффузно, захватывая большое количество разных клеток. Однако результаты прямого изучения активности отдельных нейронов ретикулярной формации оказались не соответствующими таким представлениям. При микроэлектродном отведении такой активности выяснилось, что близкорасположенные клетки могут обладать совершенно различными функциональными характеристиками. Поэтому приходится думать, что организация межнейронных связей в ретикулярной формации достаточно дифференцирована и отдельные ее клетки соединены между собой довольно специфичными связями.
2. Неспецифические нисходящие влияния
В 1946 г. американский нейрофизиолог X.Мегоун и его сотрудники обнаружили, что ретикулярная формация мозгового ствола имеет непосредственное отношение к регуляции не только вегетативной, но и соматической рефлекторной деятельности. Раздражая различные точки ретикулярной формации, можно чрезвычайно эффективно изменять течение спинальных двигательных рефлексов. В 1949 г. совместные работы X.Мегоуна и итальянского нейрофизиолога Дж.Моруцци показали, что раздражение ретикулярной формации эффективно влияет и на функции высших структур мозга, в частности коры больших полушарий, определяя переход ее в активное (бодрствующее) или неактивное (сонное) состояние. Эти работы сыграли исключительно важную роль в современной нейрофизиологии, поскольку они продемонстрировали, что ретикулярная формация занимает особое место среди других нервных центров, определяя в значительной мере общий уровень активности последних.
Влияния на двигательную деятельность спинного мозга возникают, в основном, при раздражении ретикулярной формации заднего мозга. Участки, которые создают эти эффекты, в настоящее время довольно хорошо определены, они совпадают с гигантоклеточным ядром ретикулярной формации продолговатого мозга и ретикулярным ядром моста. Эти ядра содержат большие клетки, аксоны которых образуют ретикулоспинальные пути.
Первые работы X.Мегоуна показали, что раздражение гигантоклеточного ядра вызывает ослабление в одинаковой мере всех спинальных двигательных рефлексов и сгибательных, и разгибательных. Поэтому он сделал вывод о том, что нисходящая система, берущая начало в вентрокаудальной части ретикулярной формации имеет неспецифическую тормозящую функцию. Несколько позже было обнаружено, что раздражение более дорсальных и оральных ее участков вызывало, наоборот, диффузное облегчающее действие на спинальную рефлекторную деятельность.
Микроэлектродные исследования эффектов, возникающих в нейронах спинного мозга при раздражении ретикулярной формации, действительно показали, что ретикулоспинальные влияния могут изменять передачу импульсации почти во всех рефлекторных дугах спинного мозга. Эти изменения оказываются очень глубокими и длительными, даже в том случае, когда ретикулярная формация раздражается всего несколькими стимулами, эффект в спинном мозге сохраняется сотни миллисекунд.
Одновременная активация большого количества ретикулоспинальных нейронов, которая имеет место в эксперименте с прямым раздражением ретикулярной формации и приводит к генерализованному изменению рефлекторной деятельности спинного мозга ситуация, конечно, искусственная. В естественных условиях такого глубокого сдвига этой деятельности, вероятно, не происходит; тем не менее, диффузное изменение рефлекторной возбудимости спинного мозга, несомненно, может иметь место при определенных состояниях мозга. Возможность диффузного ослабления рефлекторной возбудимости можно представить себе, например, при сне; оно приведет к снижению деятельности двигательной системы, характерному для сна. Важно учесть, что ретикулярное торможение захватывает также спинальные нейроны, участвующие в передаче афферентной импульсации в восходящем направлении, следовательно, оно должно ослаблять передачу сенсорной информации в высшие мозговые центры.
Синаптические механизмы диффузных влияний ретикулярной формации на нейроны спинного мозга изучены пока недостаточно. Как уже указывалось, эти влияния чрезвычайно длительны; кроме того, ретикулярное торможение устойчиво к действию стрихнина. Стрихнин является специфическим ядом, устраняющим постсинаптическое торможение мотонейронов, вызванное импульсацией из первичных афферентов и связанное, по всей вероятности, с выделением медиатора глицина. Нечувствительность диффузного ретикулярного торможения к стрихнину говорит, по-видимому, о том, что ретикулярные тормозящие эффекты создаются действием на спинальные клетки другого медиатора. Гистохимические исследования показали, что некоторое количество волокон в нисходящих путях, идущих из ретикулярной формации, является по своей природе адренэргическим. Однако пока неизвестно, имеют ли эти волокна отношение к диффузным ретикулоспинальным тормозящим эффектам.
Наряду с диффузными тормозящими влияниями, при раздражении определенных областей ретикулярной формации можно вызвать более специфические изменения деятельности спинальных элементов.
Если сравнить нисходящие влияния ретикулярной формации на нейронные структуры, регулирующие соматические и висцеральные функции, то можно обнаружить в них определенное сходство. Как сосудодвигательная, так и дыхательная функции ретикулярной формации строятся на сочетании деятельности двух реципрокно связанных между собой групп нейронов, оказывающих противоположное действие на спинальные структуры. Ретикулярные влияния на спинальные двигательные центры также состоят из противоположных, тормозящих и облегчающих компонентов. Поэтому создается впечатление, что реципрокный принцип организации нисходящих проекций представляет собой общее свойство ретикулярных структур; конечный эффект, соматический или вегетативный, определяется только тем, куда направлены аксоны соответствующих ретикулярных клеток. Такое сходство можно отметить и в других особенностях функционирования ретикулярных нейронов. Ретикулярные структуры, регулирующие вегетативные функции, отличаются высокой химической чувствительностью; влияния ретикулярной формации на двигательные центры также легко изменяются под влиянием таких химических факторов, как уровень CO 2 в крови и содержание в ней физиологически активных веществ (адреналина). Механизм действия адреналина на ретикулярные нейроны довольно долго вызывал споры. Дело в том, что адреналин даже при его введении непосредственно в мозговую артерию может оказывать на ретикулярные нейроны непрямое действие (путем, например, сужения мозговых сосудов с последующей аноксией мозговой ткани). Однако исследование реакций ретикулярных нейронов в ответ на прямое приложение к ним адреналина через внеклеточный микроэлектрод показало, что некоторые из них действительно являются адреноцептивными.
3. Восходящие влияния
Наряду с функциями, которые осуществляются через нисходящие пути, у ретикулярной формации есть не менее существенные функции, которые осуществляются через ее восходящие пути. Они связаны с регуляцией активности высших отделов мозга, главным образом, коры больших полушарий. Данные о том, что ретикулярная формация играет важную роль в поддержании нормальной деятельности коры больших полушарий, были получены еще в тридцатые годы нашего столетия, однако важность их в свое время не могла быть достаточно оценена. Бельгийский нейрофизиолог Ф.Бремер (1935), проводя перерезку головного мозга на различных уровнях (рис. 1) и наблюдая за функциями отделенных от остальной центральной нервной системы участков головного мозга, обратил внимание на то, что имеется чрезвычайно существенное различие между животным, у которого перерезка проведена по межколликулярному уровню (т.е. между передними и задними холмами четверохолмия), и животным, у которого линия разреза прошла между продолговатым и спинным мозгом.
Бремер сделал вывод, что для бодрствующего состояния высших отделов мозга необходимо непрерывное поступление к ним афферентной импульсации, в частности от такой обширной рецептивной зоны, как зона тройничного нерва. Предполагалось, что эта импульсация поступает в кору больших полушарий по восходящим афферентным путям и поддерживает высокую возбудимость ее нейронов.
Однако дальнейшие исследования показали, что для поддержания бодрствующего состояния коры больших полушарий важно не просто поступление к ней импульсации по афферентным системам. Если сделать перерезку ствола мозга так, чтобы не повредить основные афферентные системы (например, систему медиальной петли), но перерезать восходящие связи ретикулярной формации, то животное тем не менее впадает в сонное состояние, конечный мозг перестает активно функционировать.
Следовательно, для поддержания бодрствующего состояния конечного мозга важно, чтобы афферентная импульсация первоначально активировала ретикулярные структуры мозгового ствола. Влияния же из ретикулярных структур по восходящим путям каким то образом определяют функциональное состояние конечного мозга. Проверить такой вывод можно путем прямого раздражения ретикулярных структур. Такое раздражение через погружные электроды было проведено Дж.Моруцци и X.Мегоуном и затем воспроизводилось во многих лабораториях в условиях хронического или полухронического опыта. Оно всегда дает однозначные результаты в виде характерной поведенческой реакции животного. Если животное находится в сонном состоянии оно просыпается, у него появляется ориентировочная реакция. После прекращения раздражения животное снова возвращается в сонное состояние. Переход от сонного к бодрствующему состоянию в период раздражения ретикулярных структур отчетливо проявляется не только в поведенческих реакциях, он может быть зарегистрирован по объективным критериям деятельности коры больших полушарий, в первую очередь по изменениям ее электрической активности.
Для коры больших полушарий характерна постоянная электрическая активность (ее запись называется электрокортикограммой). Эта электрическая активность состоит из небольших по амплитуде (30-100 мкВ) колебаний, которые легко отводятся не только от открытой поверхности мозга, но и от кожи головы. У человека в спокойном дремотном состоянии такие колебания имеют частоту 8-10 в секунду и являются довольно регулярными (альфа-ритм). У высших позвоночных этот ритм менее регулярен, а частота колебаний варьирует от 6-8 у кролика до 15--20 у собаки и обезьяны. Во время активности регулярные колебания сменяются сразу же значительно меньшими по амплитуде и более высокочастотными колебаниями (бета-ритм). Появление периодических больших колебаний ясно говорит о том, что электрическая активность каких-то элементов в коре развивается синхронно. Когда правильные колебания большой амплитуды сменяются низковольтными, частыми колебаниями, то это, очевидно, говорит о том, что клеточные элементы коры начинают функционировать менее синхронно, поэтому такой тип активности называется реакцией десинхронизации. Таким образом, переход от спокойного, неактивного состояния коры к активному связан в электрическом отношении с переходом от синхронизированной активности ее клеток к десинхронизированной.
Характерным эффектом восходящих ретикулярных влияний на корковую электрическую активность является именно реакция десинхронизации. Эта реакция закономерно сопровождает описанную выше поведенческую реакцию, характерную для ретикулярных влияний. Реакция десинхронизации не ограничивается каким-нибудь одним участком коры, а регистрируется от больших ее областей. Это говорит о том, что восходящие ретикулярные влияния являются генерализованными.
Описанные изменения в электроэнцефалограмме не являются единственным электрическим проявлением восходящих ретикулярных влияний. В определенных условиях можно выявить более прямые эффекты ретикулярной импульсации, поступающей к коре больших полушарий. Они впервые были описаны в 1940 г. американскими исследователями А.Форбсом и Б.Мориссоном, которые исследовали вызванную электрическую активность коры при различных афферентных влияниях. При раздражении какой-либо афферентной системы в соответствующей проекционной зоне коры обнаруживается электрический ответ, указывающий на приход к этой области афферентной волны, этот ответ получил название первичного ответа. Кроме этого локального ответа, афферентное раздражение вызывает длиннолатентный ответ, возникающий в обширных областях коры мозга. Этот ответ Форбс и Мориссон назвали вторичным ответом.
То обстоятельство, что вторичные ответы возникают со скрытым периодом, значительно превышающим скрытый период первичного ответа, ясно говорит о том, что они связаны с поступлением афферентной волны в кору не по прямым, а по каким-то окольным связям, через дополнительные синаптические переключения. Позже, когда было применено прямое раздражение ретикулярной формации, было показано, что оно может вызвать ответ такого же типа. Это позволяет заключить, что вторичный ответ является электрическим проявлением поступления в кору больших полушарий афферентное волны через ретикулокортикальные связи.
Через ствол мозга проходят прямые афферентные пути, которые после синаптического перерыва в таламусе поступают в кору больших полушарий. Приходящая по ним афферентная волна вызывает первичный электрический ответ с соответствующей проекционной зоне с коротким скрытым периодом. Одновременно афферентная волна по коллатералям ответвляется в ретикулярную формацию и активирует ее нейроны. Затем по восходящим путям от нейронов ретикулярной формации импульсация тоже поступает в кору, но уже в виде задержанной реакции, возникающей с большим скрытым периодом. Эта реакция охватывает не только проекционную зону, но и большие участки коры, вызывая в них какие-то изменения, важные для бодрствующего состояния.
Нисходящие функции ретикулярной формации включают в себя, как правило, облегчающие и тормозящие компоненты, которые осуществляются по реципрокному принципу. Швейцарский физиолог В. Гесс (1929) впервые показал, что в стволе мозга можно найти точки, при раздражении которых у животного наступает сон. Гесс назвал эти точки центрами сна. Позже Дж.Моруцци и сотр. (1941) также обнаружили, что, раздражая некоторые участки ретикулярной формации заднего мозга, можно у животных вызвать в коре вместо десинхронизации синхронизацию электрических колебаний н соответственно перевести животное из бодрствующего состояния в пассивное, сонное. Поэтому можно думать, что в составе восходящих путей ретикулярной формации действительно существуют не только активирующие, но и инактивирующие подразделения, последние каким-то образом снижают возбудимость нейронов конечного мозга.
Нейронная организация восходящей системы ретикулярной формации не вполне ясна. При разрушении ретикулярных структур среднего и заднего мозга в коре больших полушарий не обнаруживается существенного количества дегенерирующих окончаний, которые могли бы быть отнесены к прямым ретикулярным волокнам. Значительная дегенерация окончаний в коре возникает только при разрушении неспецифических ядер таламуса. Поэтому возможно, что восходящие ретикулярные влияния передаются в кору больших полушарий не по прямым путям, а через какие-то промежуточные синаптические соединения, локализованные, вероятно, в промежуточном мозге.
Любопытно отметить, что гистологические и электрофизиологические данные указывают на характерную деталь хода аксонов многих ретикулярных нейронов. Аксоны нейронов гигантоклеточного ядра, т. е. основного ядра ретикулярной формации, очень часто делятся Т-образно, и один из отростков идет вниз, образуя ретикулоспинальный путь, а второй вверх, направляясь в верхние отделы головного мозга. Создается такое впечатление, что и восходящие, и нисходящие функции ретикулярной формации могут быть связаны с деятельностью одних и тех же нейронов. По функциональным свойствам ретикулярные структуры, создающие восходящие влияния, также имеют много общего со структурами, обеспечивающими нисходящие влияния. Восходящие влияния несомненно носят тонический характер, они легко изменяются гуморальными факторами и высокочувствительны к фармакологическим веществам. Снотворное и наркотическое действие барбитуратов основано, по-видимому, именно на блокировании в первую очередь восходящих влияний ретикулярной формации.
4. Свойства ретикулярных нейронов
Большой интерес представляют результаты исследований общих закономерностей деятельности нейронов ретикулярной формации. Этим исследованиям положили начало работы Дж.Моруцци, который впервые подробно описал функциональные свойства ретикулярных нейронов, использовав при этом внеклеточное микроэлектродное отведение. При этом сразу же обратила на себя внимание их способность к стойкой импульсной активности. Если деятельность ретикулярной формации исследуется в условиях отсутствия наркоза, то большинство ее нейронов постоянно генерирует нервные разряды с частотой порядка 5-10 в секунду. Различные афферентные влияния суммируются с этой фоновой активностью ретикулярных нейронов, вызывая в одних из них ее учащение, а в других торможение. Причина постоянной фоновой активности в этих условиях, по-видимому, двоякая. С одной стороны, она может быть связана с высокой химической чувствительностью мембраны ретикулярных клеток и постоянной ее деполяризацией гуморальными химическими факторами. С другой стороны, она может определяться особенностями афферентных связей ретикулярных нейронов, а именно, конвергенцией к ним коллатералей от огромного количества различных сенсорных путей,
поэтому даже в случае, когда организм не подвергается действию каких-нибудь специальных раздражителей в ретикулярную формацию могут непрерывно поступать импульсы от всевозможных неконтролируемых воздействий (Рис. 2). Эти импульсы, приходя к клеткам через многочисленные синаптические входы, вызывают также дополнительную деполяризацию мембраны. В связи с таким характером активности ретикулярных клеток их влияния на другие структуры также носят постоянный, тонический характер. Если, например, искусственно прервать связь ретикулярной формации со спинным мозгом, то это сразу же вызовет существенные стойкие изменения рефлекторной деятельности последнего; в частности, облегчаются рефлексы, осуществляющиеся по полисинаптическим путям. Это ясно говорит о том, что спинальные нервные элементы находятся под тоническим, преимущественно тормозящим. контролем со стороны ретикулярной формации.
При изучении деятельности отдельных нейронов ретикулярной формации обнаруживается еще одно очень существенное их свойство. Оно вытекает из высокой химической чувствительности ретикулярных нейронов и проявляется в легком блокировании их деятельности фармакологическими веществами. Особенно активными оказываются соединения барбитуровой кислоты, которые даже в небольших концентрациях, не действующих на спинальные нейроны или на нейроны коры больших полушарий, полностью прекращают активность ретикулярных нейронов. Эти соединения очень легко связываются с хемоцептивными группировками мембраны последних.
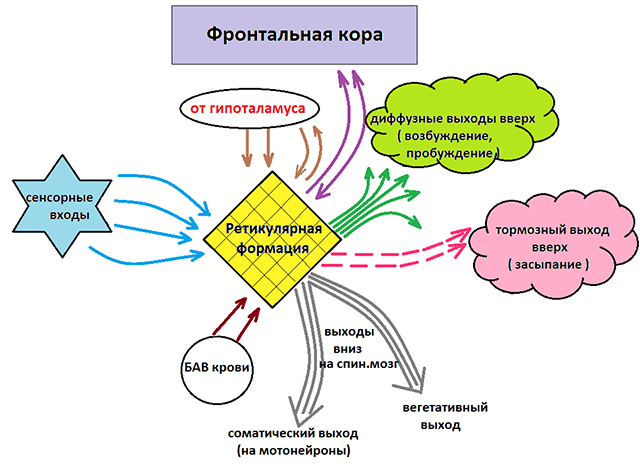
· Ретикулярная формация - это продолговатая структура в стволе мозга. Она представляет собой важный пункт на пути восходящей неспецифической соматосенсорной чувствительности. К ретикулярной формации приходят также пути от всех других афферентных черепных нервов, т.е. практически от всех органов чувств. Дополнительная афферентация поступает от многих других отделов головного мозга - от моторных областей коры и сенсорных областей коры, от таламуса и гипоталамуса.
· Имеется также множество эфферентных связей нисходящие к спинному мозгу и восходящие через неспецифические таламические ядра к коре головного мозга, гипоталамусу и лимбической системе.
· Большинство нейронов образует синапсы с двумя - тремя афферентами разного происхождения, такая полисенсорная конвергенция характерна для нейронов ретикулярной формации.
· Другими их свойствами являются большие рецептивные поля поверхности тела, часто билатеральные, длительный латентный период ответа на периферическую стимуляцию (вследствие мультисинаптического проведения), слабая воспроизводимость реакции.
· Функции ретикулярной формации изучены не полностью. Считается, что она участвует в следующих процессах:
в регуляция уровня сознания путем воздействия на активность корковых нейронов, например, участие в цикле сон – бодорствование.
в придании аффективно-эмоциональной окраски сенсорным стимулам, в том числе болевым сигналам, идущим по переднебоковому канатику, путем проведения афферентной информации к лимбической системе.
в вегетативных регулирующих функциях, в том числе во многих жизненно важных рефлексах, при которых должны взаимно координироваться разные афферентные и эфферентные системы.
в позных и целенаправленных движениях в качестве важного компонента двигательных центров ствола мозга.
Список литературы
1. «Анатомия человека» в двух томах под ред. М.Р. Сапина М. «Медицина» 1986 г.
2. М.Г. Привес, Н.К. Лысенков, В.И. Бушкович «Анатомия человека» М. «Медицина» 1985 г.
3. Костюк П.Г. «Ретикулярная формация мозгового ствола»
4. Schmidt R.F., Thews G., “Human Physiology”, 1983
Наряду с быстро реагирующей на раздражители первой активизирующей системой, в которую входят проводящие пути, существует еще и неспецифическая система замедленного ответа на внешние импульсы, которая является филогенетически более древней, чем остальные структуры мозга и напоминает диффузный тип нервной системы. Эта структура называется ретикулярной формацией (РФ) и состоит из более чем 100 ядер, соединенных между собой. РФ простирается от ядер таламуса и субталамуса до промежуточной зоны спинного мозга верхних шейных сегментов.
Первые описания РФ были сделаны немецкими морфологами: в 1861 г. К. Рейхертом и в 1863 г. О. Дейтерсом, который и ввел термин РФ; большой вклад в ее изучение внес В.М. Бехтерев.
Нейроны, составляющие РФ разнообразны по величине, строению и функции; имеют обширно разветвленное дендритное дерево и длинные аксоны; их отростки густо переплетаются, напоминая сеть (лат. reticulum - сеточка, formatio - образование).
Свойства ретикулярных нейронов :
1. Мультипликация (умножение импульса) и амплификация (получение конечного большого результата) – осуществляется благодаря сложному переплетению отростков нейронов. Поступающий импульс многократно умножается, что в восходящем направлении дает ощущение даже небольших стимулов, а в нисходящем направлении (ретикулоспинальные пути) позволяет вовлекать в ответную реакцию множество структур НС.
2. Генерирование импульсов . Д. Моруцци доказал, что большинство нейронов РФ постоянно генерирует нервные разряды с частотой порядка 5-10 в секунду. Различные афферентные стимулы суммируются с этой фоновой активностью ретикулярных нейронов, вызывая в одних из них ее учащение, а в других торможение.
3. Полисенсорность . Почти все нейроны РФ способны отвечать на на раздражения самых различных рецепторов. Тем не менее, часть из них реагируют на кожные раздражения и на свет, другие на звуковые и кожные раздражения и т. д. Таким образом, полного смешивания афферентных сигналов в ретикулярных нейронах не происходит; в их связях есть частичная внутренняя дифференциация.
4. Чувствительность к гуморальным факторам и, особенно, к фармпрепаратам. Особенно активными оказываются соединения барбитуровой кислоты, которая даже в небольших концентрациях полностью прекращают активность ретикулярных нейронов, при этом не действуя на спинальные нейроны или на нейроны коры больших полушарий.
В целом же, РФ характеризуется разлитыми рецептивными полями, длительным латентным периодом ответа на периферическую стимуляцию и слабой воспроизводимостью реакции.
Классификация :
Существует топографическая и функциональная класификация РФ.
I. Топографически вся ретикулярная формация может быть подразделена на каудальный и ростральный отделы.
1. Ростральные ядра (ядра среднего мозга и верхней части моста, связанные с промежуточным мозгом) – отвечают за состояние возбуждения, бодрствования, настороженности. Ростральные ядра оказывают локальное влияние на определенные зоны коры больших полушарий. Поражение этого отдела вызывает сонливость.
2. Каудальные ядра (моста и промежуточного мозга, связаны с ядрами черепных нервов и спинным мозгом) – выполняют моторную, рефлекторную и вегетативную функции. Некоторые ядра в процессе эволюции получили специализацию - вазомоторный центр (депрессорные и прессорные зоны), дыхательный центр (экспираторный и инспираторный), рвотный центр. Каудальный отдел РФ оказывает более разлитое, генерализованное влияние на обширные зоны головного мозга. Поражение этого отдела вызывает бессонницу.
Если рассматривать ядра РФ каждого отдела мозга, то РФ таламуса образуют капсулу латерально вокруг зрительных бугров. Они получают импульсы от коры и дорзальных ядер таламуса. Функция ретикулярных ядер таламуса – фильтрация сигналов, идущих через таламус к коре полушарий; проекция их на другие ядра таламуса. В целом, они влияют на всю поступающую сенсорную и когнитивную информацию.
Ядра РФ среднего мозга включают покрышечные ядра: nuclei tegmentalis dorsalis et ventralis , nucleus cuneiformis . Они получают импульсы через fasciculus mammillo-tegmentalis (Gudden) , который является частью мамиллоталямического пути.
РФ моста, образованная околосрединными (парамедианными) ядрами, не имеет четких границ. Эти ядра участвуют в скоординированном движении глаз, фиксированном взгляде и саккадном движении глаз (быстрые синхронные движения глаз). РФ моста лежит кпереди и латерально от медиального продольного пучка, получает импульсы по нервным волокнам от верхних бугорков четверохолмия через предорзальные нервные волокна и от передних зрительных полей посредством лобномостовых связей.
Латеральную РФ образуют преимущественно ядра РФ продолговатого мозга. Эта структура имеет множество ганглиев, вставочных нейронов вокруг черепных нервов, которые служат для модулирования связанных с ними рефлексов и функций.
II. Функционально ядра РФ подразделяются на вертикальные образования:
1. Срединный столб (ядра шва) – узкий парный столб клеток вдоль срединной линии ствола мозга. Простираются от продолговатого до среднего мозга. Дорзальные ядра шва синтезируют серотонин.
2. Медиальный столб (голубоватое пятно) – относится к РФ. Клетки голубоватого пятна синтезируют норэпинефрин, аксоны идут к областям коры, ответственным за возбуждение (бодрствование).
3. Латеральный столб (серое вещество вокруг Сильвиева водопровода) – (входит в состав лимбической системы) – клетки имеют опиоидные рецепторы, что способствует эффекту обезболивания.
Функция РФ:
1. Регуляция сознания путем изменения активности корковых нейронов, участие в цикле сон /бодрствование, возбуждение, внимание, обучаемость - когнитивные функции
2. Обеспечение эмоциональной окраски сенсорным стимулам (ретикулолимбические связи)
3. Участие в жизненно важных вегетативных реакциях (сосудодвигательный, дыхательный, кашлевой, рвотный центры)
4. Реакция на боль – РФ проводит болевые импульсы к коре и образует нисходящие аналгезирующие пути (влияет на спинной мозг, частично блокируя передачу болевых импульсов от спинного мозга к коре)
5. Привыкание – это процесс, в котором мозг учится игнорировать незначительные повторяющиеся стимулы извне, отдавая предпочтение новым стимулам. Пример – способность спать в переполненном шумном транспорте, сохраняя способность проснуться на сигнал автомобиля или крик ребенка
6. Соматодвигательный контроль – обеспечивается ретикулоспинальными путями. Эти пути отвечают за мышечный тонус, равновесие, положение тела в пространстве, особенно при его движении
7. Формирование интегрированных реакций организма на стимулы, например, сочетанная работа речедвигательного аппарата, обшая двигательная активность.
Связи РФ
Аксоны РФ связывают практически все структуры мозга между собой. РФ морфологически и функционально связана со спинным мозгом, мозжечком, лимбической системой и корой больших полушарий головного мозга.
Часть аксонов РФ имеет нисходящее направление и образует ретикулоспинальные пути, а часть восходящее (спиноретикулярные пути). Возможна также циркуляция импульсов по замкнутым нейронным цепям. Таким образом, существует постоянный уровень возбуждения нейронов РФ, вследствие чего обеспечиваются тонус и определённая степень готовности к деятельности различных отделов центральной нервной системы. Степень возбуждения РФ регулируется корой больших полушарий головного мозга.
1. Спиноретикулярные (спиноретикулокортикальные) пути (восходящая активизирующая ретикулярная система) - получают импульсы от аксонов восходящих (сенсорных) путей общей и специальной чувствительности. Соматовисцеральные волокна идут в составе спиноретикулярного тракта (переднебоковой канатик), а также в составе проприоспинальных путей и соответствующих путей от ядра спинального тройничного тракта. К ретикулярной формации приходят также пути от всех других афферентных черепномозговых нервов, т.е. практически от всех органов чувств. Дополнительная афферентация поступает от многих других отделов головного мозга - от моторных областей коры и сенсорных областей коры, мозжечка, базальных ядер, красного ядра, от таламуса и гипоталамуса. Эта часть РФ отвечает за процессы возбуждения, внимания, бодрствования, обеспечивает эмоциональные реакции, важные при познавательном процессе. Поражения, опухоли этой части РФ вызывают снижение уровня сознания, активности психической деятельности, в частности когнитивных функций, двигательной активности, синдром хронической усталости. Возможны сонливость, проявления оглушенности, общей и речевой гипокинезии, акинетического мутизма, сопора, в тяжелых случаях - кома.
2. Ретикулоспинальные пути (нисходящие ретикулярные связи) – могут оказывать как стимулирующее влияние (отвечают за мышечный тонус, вегетативные функции, активирует восходящую РФ), так и угнетающее (способствуют плавности и точности произвольных движений, регулирует мышечный тонус, положение тела в пространстве, вегетативные функции, рефлексы). Обеспечиваются множеством эфферентных связей - нисходящие к спинному мозгу и восходящие через неспецифические таламические ядра к коре головного мозга, гипоталамусу и лимбической системе. Большинство нейронов образует синапсы с двумя - тремя дендритами разного происхождения, такая полисенсорная конвергенция характерна для нейронов ретикулярной формации.
3. Ретикуло-ретикулярные связи .
